

- Syncytiotrophoblasts (STBs): Multinucleated cells formed through CTB fusion that mediate nutrient and gas exchange, hormone production, and immune regulation at the maternal–fetal interface.
- Extravillous trophoblasts (EVTs): Invasive trophoblasts that migrate into the maternal decidua and remodel uterine spiral arteries to establish low-resistance blood flow to the developing fetus.
- Reliance on collagen IV coated plastic cultureware increases cost and technical complexity.
- Atmospheric oxygen conditions accelerate senescence and spontaneous differentiation.
- Hypoxic culture conditions (2% O₂), while partially protective, require specialized infrastructure that limits accessibility and reproducibility.
- CTBs rapidly lose progenitor-associated phenotypes during serial passaging.
- Freshly isolated CTBs from term placentas have significant donor-to-donor variability and some cell populations are nearly senescent when compared to early term placentas.
- Standard 2D systems fail to reproduce the three-dimensional microenvironment and biomechanical cues that regulate trophoblast organization and lineage specification (Fig. 1).
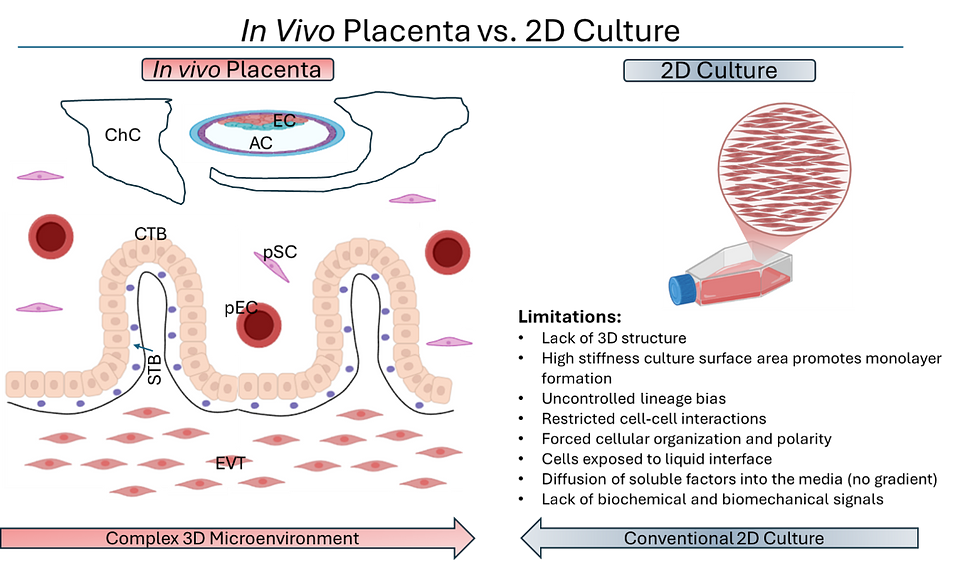
Figure 1: In vivo placental structure versus conventional 2D culture limitations. Comparison of the complex placental microenvironment vs 2D culture. Image on the left adapted and simplified from Knofler et al., 2019. AC amniotic cavity, ChC chorionic cavity, CTB cytotrophoblast, EC ectoderm, EVT extra-villous trophoblast, pSC placental stromal cell, STB syncytiotrophoblasts.
- Bio-Blocks® hardware: Tissue-mimetic hydrogel substrates.
- Standardized process logic and SOPs.
- The living biology cultured within the system.

Figure 2. Bio-OS: Ronawk’s Biological Operating System, a novel platform-based technology for the culture of cells in a physiologically relevant tissue-mimetic environment.
- Efficient nutrient and oxygen diffusion.
- Continuous cell migration and expansion.
- Eliminates need for repeated subculturing.
- Enhanced retention of stem-like phenotypes.
- Retention of differentiation potential.
- Easy collection and enhanced secretion of biologics including extracellular vesicles, proteins, and antibodies
- Together, these features reduce cellular stress, minimize phenotype drift and contamination risk, and deliver reproducible, high-fidelity tissue models that scale seamlessly from discovery to production within a compact footprint.
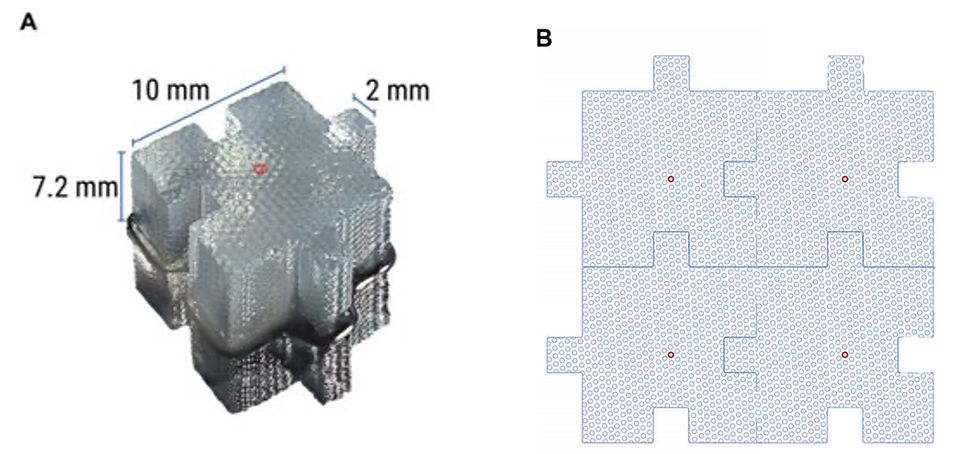
Figure 3. Schematic of Bio-Block architecture and porous microchannel structure. (A) Classic Bio-Block in the puzzle piece configuration. Bio-Blocks are ~1-cm3 and have the equivalent culturable surface area as a T150 flask. Center microchannel shown with red circle. (B) Illustrative depiction of interconnected microchannels.
| 2D | Bio-Block |
Seeding Density | 750,000 | 500,000* |
| Culture Surface Area | 75cm2 | ~150cm2 |
CTB passage at time start of experiment | 3 | 3 |
Number of subpassages | 5 | 0 |
Total days in culture | 20 | 20 |
Temperature | 37°C | 37°C |
Oxygen | Atmospheric | Atmospheric |
CO2 | 5% | 5% |
Vessel coating | 1% Gelatin | Proprietary hydrogel polymer |
Number of Bio-Blocks per well | N/A | 4 (12 total) |
*Seeding density adjusted for total number of cells available.
- DAPI/Hoechst 33342 (DNA)
- Wheat Germ Agglutinin (Lectins)
- Phalloidin (F-actin)
- MemGlow (Lipids)
- CD49f: Cytotrophoblast progenitor marker.
- CK7: Epithelial trophoblast marker.
- CD138: Syncytiotrophoblast-associated marker.
- HLA-G: Sxtravillous trophoblast marker.
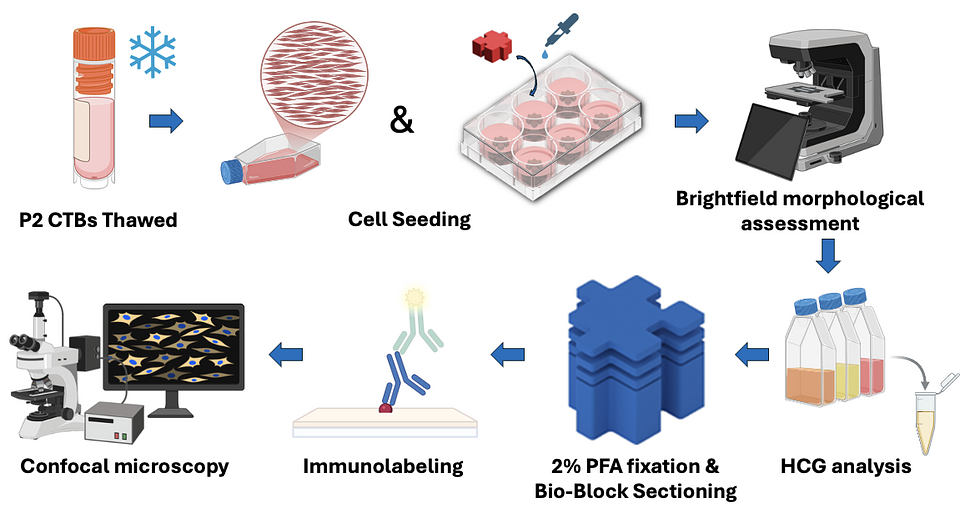
Figure 4. Experimental workflow schematic. Image created using Bio-Render.
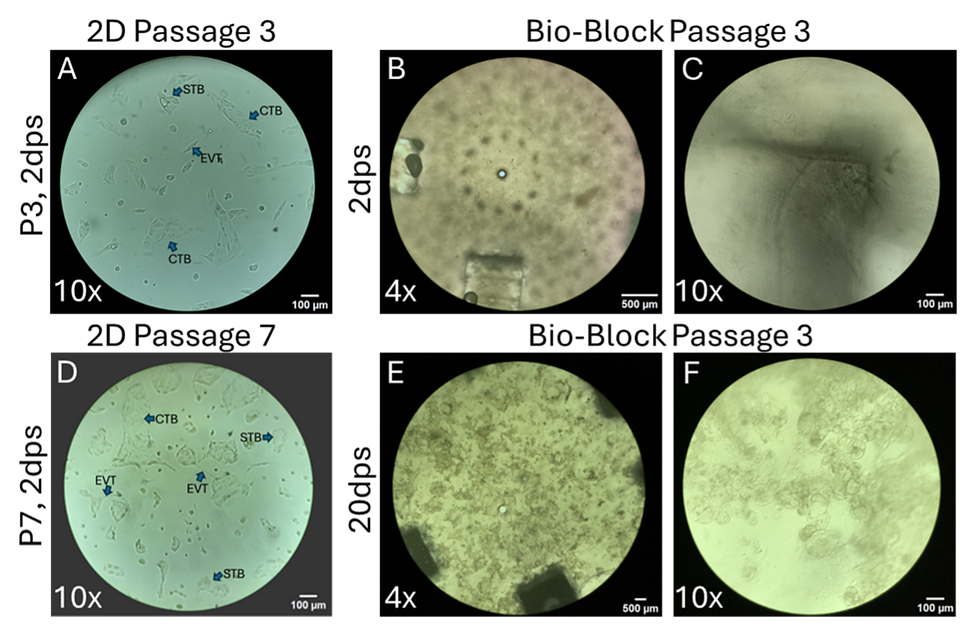
Figure 5: Brightfield Morphological Assessment of 2D and 3D Cultures. (A, D) show representative images of CTBs cultured on gelatin coated T75 flasks and serial passaged over 20 days. B, C, E, F) show CTBs cultured in Ronawk 500V Bio-Blocks for 20 days without subculturing. dps, days post seed.
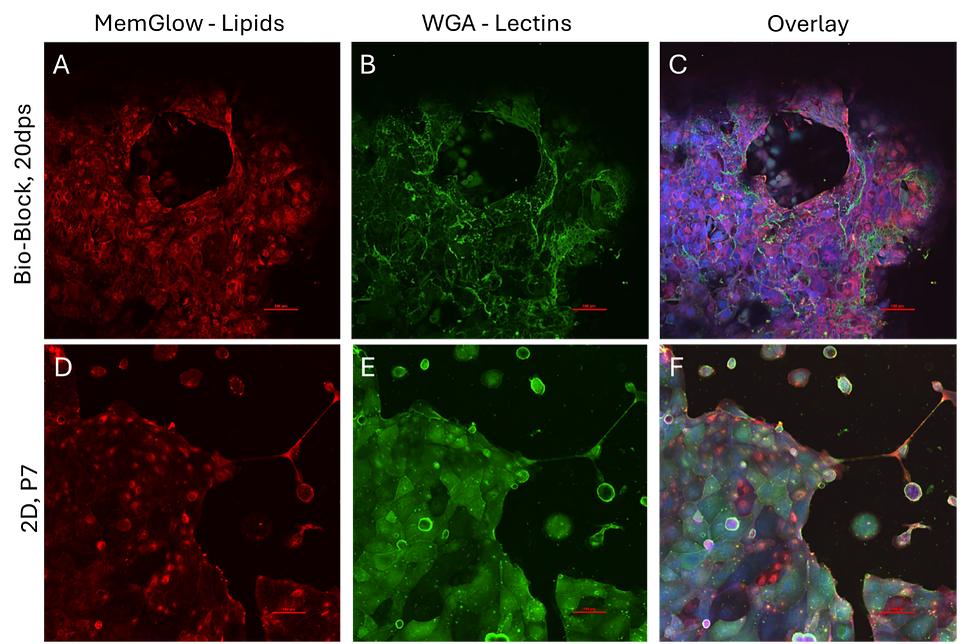
Figure 6: Morphological assessment panels. (A, B, C) CTBs cultured in Ronawk Bio-Block for 20 days followed by PFA fixation and morphological staining with Memglow (A), WGA (B), DAPI, and Phalloidin. 3D overlay (C). (D, E, F) Passage 7 CTBs cultured on 24-well gelatin coated glass bottom plate, fixed in 2% PFA and stained with morphological markers Memglow (D), WGA (E), DAPI and Phalloidin. 2D overlay (F).
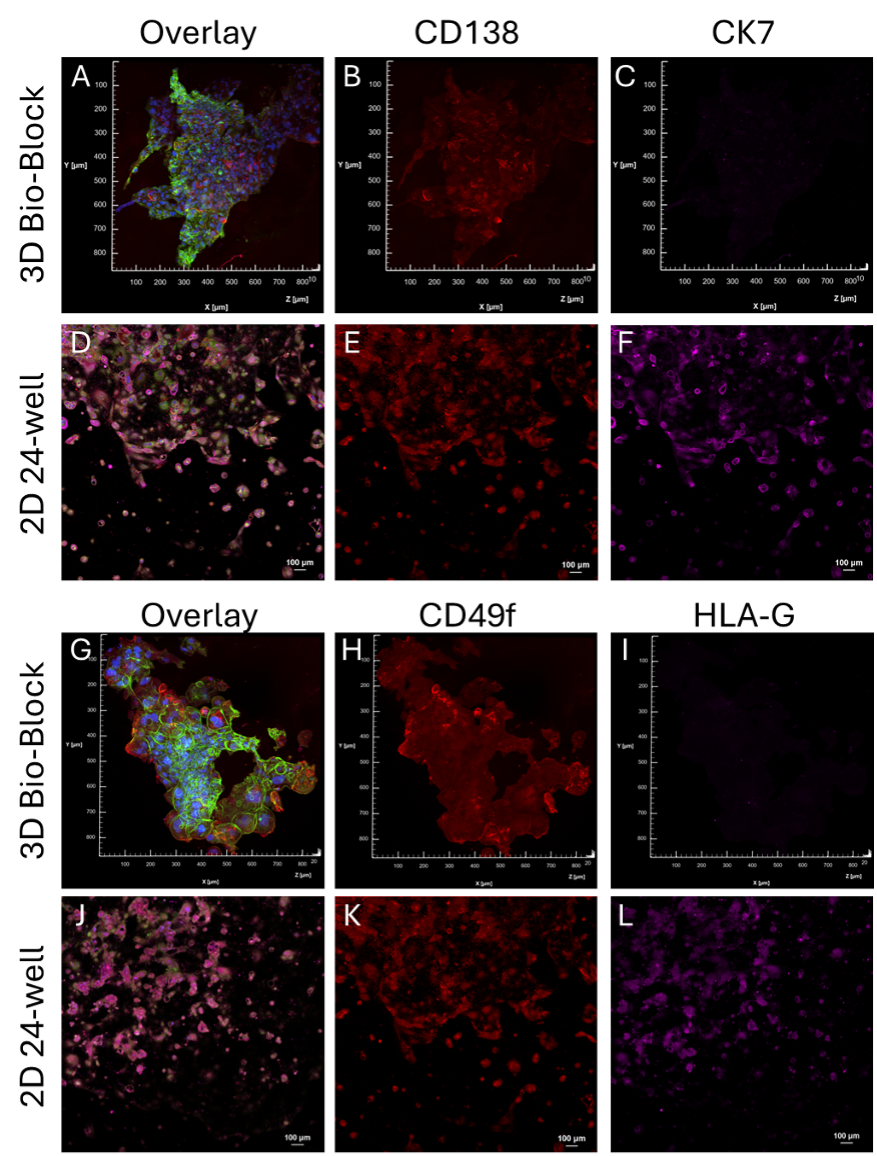
Figure 7. Immunofluorescence staining panels: Evaluation of CTB lineage markers CTBs cultured in Bio-OS or 2D. Blue, Hoechst, Green WGA, Red CD138 (A, B, D, E) or CD49f (G, H, J, K), Magenta CK7 (A, C,D, F) or HLAG (G, I, J, L). 3D max-project of 50 µM Bio-Block section overlay (A,G). P8 2D culture overlay (D, J). Fluorescent threshold of the 2D panels for CD138 and CD49f was increased for easier visualization.
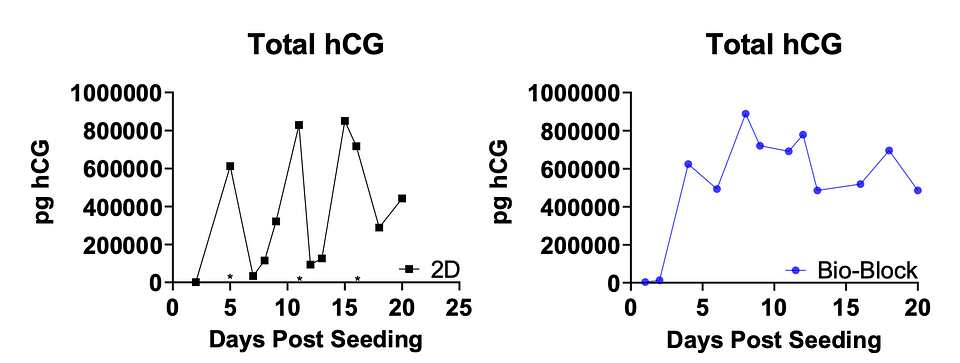
Figure 8: Longitudinal hCG expression. 3D Bio-Blocks shown in Blue, 2D shown in Black. *subculturing of 2D cells.
Feature | 2D Culture | Bio-OS Platform |
Structure | Monolayer | 3D organization |
Phenotype retention | Poor over passage | Enhanced |
Differentiation bias | STB skewed | Balanced potential |
Culture duration | Requires passaging | Long-term without passaging |
Figure 9. Potential applications of Bio-OS in placental biology
- Follow Ronawk on LinkedIn to stay updated on our latest platform advancements, clinical milestones, and partnership announcements.
- Share this white paper with your network, colleagues, or investment committees who are looking for the next paradigm shift in the life sciences.
About Ronawk
Ronawk Media Contact
- Knöfler, M. et al. Human placenta and trophoblast development: key molecular mechanisms and model systems. Cell. Mol. Life Sci. 76, 3479–3496 (2019).
- Li, X., Li, Z.-H., Wang, Y.-X. & Liu, T.-H. A comprehensive review of human trophoblast fusion models: recent developments and challenges. Cell Death Discov. 9, 372 (2023).
- Hawkins, A. et al. Long-term culture and passaging of term trophoblast for the investigation of syncytiotrophoblast function. Placenta 166, 25–32 (2025).
- Haider, S. et al. Notch1 controls development of the extravillous trophoblast lineage in the human placenta. Proc. Natl. Acad. Sci. U.S.A. 113, (2016).
- Cindrova-Davies, T. & Sferruzzi-Perri, A. N. Human placental development and function. Seminars in Cell & Developmental Biology 131, 66–77 (2022).
- Brosens, J. J., Pijnenborg, R. & Brosens, I. A. The myometrial junctional zone spiral arteries in normal and abnormal pregnancies. American Journal of Obstetrics and Gynecology 187, 1416–1423 (2002).
- Khong, T. Y., De Wolf, F., Robertson, W. B. & Brosens, I. Inadequate maternal vascular response to placentation in pregnancies complicated by pre‐eclampsia and by small‐for‐gestational age infants. BJOG 93, 1049–1059 (1986).
- Loregger, T., Pollheimer, J. & Knöfler, M. Regulatory Transcription Factors Controlling Function and Differentiation of Human Trophoblast—A Review. Placenta 24, S104–S110 (2003).
- Murthi, P. et al. Homeobox genes and down-stream transcription factor PPARγ in normal and pathological human placental development. Placenta 34, 299–309 (2013).
- Sonderegger, S., Pollheimer, J. & Knöfler, M. Wnt Signalling in Implantation, Decidualisation and Placental Differentiation – Review. Placenta 31, 839–847 (2010).
- Liu, L., Tang, L., Chen, S., Zheng, L. & Ma, X. Decoding the molecular pathways governing trophoblast migration and placental development; a literature review. Front. Endocrinol. 15, 1486608 (2024).
- Li, L. & Schust, D. J. Isolation, purification and in vitro differentiation of cytotrophoblast cells from human term placenta. Reprod Biol Endocrinol 13, 71 (2015).
- Merck. Evolution of Cell Culture Surfaces. https://www.merckmillipore.com/VE/en/technical-documents/technical-article/cell-culture-and-cell-culture-analysis/mammalian-cell-culture/evolution-of-cell.
- Bissell, M. J., Hall, H. G. & Parry, G. How does the extracellular matrix direct gene expression? Journal of Theoretical Biology 99, 31–68 (1982).
- Knight, E. & Przyborski, S. Advances in 3D cell culture technologies enabling tissue‐like structures to be created in vitro. Journal of Anatomy 227, 746–756 (2015).
- Li, N., Tourovskaia, A. & Folch, A. Biology on a Chip: Microfabrication for Studying the Behavior of Cultured Cells. Crit Rev Biomed Eng 31, 423–488 (2003).
- Bissell, M. J. & Radisky, D. Putting tumours in context. Nat Rev Cancer 1, 46–54 (2001).
- Serban, M. A. & Prestwich, G. D. Modular extracellular matrices: Solutions for the puzzle. Methods 45, 93–98 (2008).
- Pampaloni, F., Reynaud, E. G. & Stelzer, E. H. K. The third dimension bridges the gap between cell culture and live tissue. Nat Rev Mol Cell Biol 8, 839–845 (2007).
- Hodge, J. G., Decker, H. E., Robinson, J. L. & Mellott, A. J. Tissue‐mimetic culture enhances mesenchymal stem cell secretome capacity to improve regenerative activity of keratinocytes and fibroblasts in vitro. Wound Repair Regeneration 31, 367–383 (2023).
- Humphrey, J. D., Dufresne, E. R. & Schwartz, M. A. Mechanotransduction and extracellular matrix homeostasis. Nat Rev Mol Cell Biol 15, 802–812 (2014).
- Dupont, S. Role of YAP/TAZ in cell-matrix adhesion-mediated signalling and mechanotransduction. Experimental Cell Research 343, 42–53 (2016).
- Engler, A. J., Sen, S., Sweeney, H. L. & Discher, D. E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 126, 677–689 (2006).
- Pittenger, M. F. et al. Mesenchymal stem cell perspective: cell biology to clinical progress. npj Regen Med 4, 22 (2019).
- Drela, K., Stanaszek, L., Nowakowski, A., Kuczynska, Z. & Lukomska, B. Experimental Strategies of Mesenchymal Stem Cell Propagation: Adverse Events and Potential Risk of Functional Changes. Stem Cells International 2019, 1–10 (2019).
- Bonab, M. M. et al. Aging of mesenchymal stem cell in vitro. BMC Cell Biol 7, 14 (2006).
- Jiang, T. et al. In vitro expansion impaired the stemness of early passage mesenchymal stem cells for treatment of cartilage defects. Cell Death Dis 8, e2851–e2851 (2017).
- Jensen, C. & Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 7, 33 (2020).
- Yin, Q. et al. Comparison of senescence-related changes between three- and two-dimensional cultured adipose-derived mesenchymal stem cells. Stem Cell Res Ther 11, 226 (2020).
- Duval, K. et al. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology 32, 266–277 (2017).
- Baker, B. M. & Chen, C. S. Deconstructing the third dimension – how 3D culture microenvironments alter cellular cues. Journal of Cell Science jcs.079509 (2012) doi:10.1242/jcs.079509.
- Tam, R. Y., Smith, L. J. & Shoichet, M. S. Engineering Cellular Microenvironments with Photo- and Enzymatically Responsive Hydrogels: Toward Biomimetic 3D Cell Culture Models. Acc. Chem. Res. 50, 703–713 (2017).
- Sheridan, M. A. et al. Establishment and differentiation of long-term trophoblast organoid cultures from the human placenta. Nat Protoc 15, 3441–3463 (2020).
- Betz D, Fane K. Human Chorionic Gonadotropin. [Updated 2025 Apr 27]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2026 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK532950/
- Massin, N. et al. Defect of Syncytiotrophoblast Formation and Human Chorionic Gonadotropin Expression in Down’s Syndrome. Placenta 22, S93–S97 (2001).
- 1. Nwabuobi, C. et al. hCG: Biological Functions and Clinical Applications. IJMS 18, 2037 (2017).